Artículo de Investigación
Inmunogenicidad y eficacia de SOBERANA® 02 contra el SARS-CoV-2 en el modelo animal hámster sirio dorado
Immunogenicity and efficacy of SOBERANA® 02 against SARS-CoV-2, in the Syrian golden hamster animal model
Nibaldo Luis González Sosa1
https://orcid.org/0000-0002-8665-4413
Reynaldo
Oliva Hernández2
https://orcid.org/0000-0001-8198-9161
Madeline
Blanco de Armas1
https://orcid.org/0000-0002-9243-066X
Mildrey
Fariñas Medina2
https://orcid.org/0000-0001-6530-9904
Juan
Francisco Infante Bouzac2
https://orcid.org/0000-0002-6369-8608
Yanet
Climent Ruiz2
https://orcid.org/0000-0002-2824-6374
Sonsire
Fernández Castillo2
https://orcid.org/0000-0001-5329-5971
Liuber
Yans Machado Zaldívar1
https://orcid.org/0000-0001-5309-8062
Enrique
Noa Romero1
https://orcid.org/0000-0003-2656-0228
Yuliet
Sotes Sarguero1
https://orcid.org/0000-0002-5014-8623
Darcy
Núñez Martínez2
https://orcid.org/0009-0007-8157-9884
Sandra
Rodríguez Salgueiro3
https://orcid.org/0000-0003-3341-128X
Mireida
Rodríguez Acosta1
https://orcid.org/0000-0002-1574-6951
Yisabel
Aranguren Mazorra2
https://orcid.org/0009-0001-1758-938X
Otto
Cruz Sui1
https://orcid.org/0000-0003-4029-4253
Marta
Dubed Echevarría1
https://orcid.org/0000-0002-0072-8590
Dagmar
García Rivera2
https://orcid.org/0000-0002-2099-1791
1Centro
de Investigaciones Científicas de la Defensa Civil (CICDC). Mayabeque, Cuba.
2Instituto
Finlay de Vacunas. La Habana, Cuba.
3Centro
de Productos Naturales. La Habana, Cuba.
Introducción:
En los estudios preclínicos de la vacuna SOBERANA® 02, fue esencial verificar
la eficacia de la respuesta inmune contra el SARS-CoV-2, en modelos animales.
En un ensayo de desafío se exponen al virus, de manera controlada, animales
inmunizados con el antígeno vacunal.
Objetivos: Evaluar la inmunogenicidad y protección del antígeno FR02
contra el SARS-CoV-2, en el modelo hámster sirio dorado.
Métodos: Los hámsteres inmunizados se retaron con la variante viral D614G.
Se controló peso corporal, signos respiratorios, presencia de ARN viral en nasofaringe
y tejido pulmonar. Se realizaron cultivos en células Vero E6 y análisis histopatológicos
de los pulmones. Se utilizaron como controles, animales infectados sin vacunar,
y animales no vacunados ni infectados.
Resultados: Los hámsteres inmunizados no presentaron signos de enfermedad;
tuvieron concentraciones de ARN viral en nasofaringe y pulmones inferiores a
los no vacunados infectados, con reducción de la carga viral (6 logaritmos)
en los pulmones, en el día del pico de replicación viral, además de no ser detectable
en un 27,7 %. Se observó efecto citopático en el 5,8 % de los cultivos virales
del pulmón de los animales inmunizados vs. los no inmunizados infectados, en
los cuales ocurrió en el 100 %. Los hámsteres inmunizados presentaron menores
índices de daño pulmonar agudo y daño histológico global.
Conclusiones:
La respuesta inmune generada en los animales inmunizados confirió protección
contra la infección, el daño pulmonar grave y el desarrollo de la enfermedad
sintomática en los hámsteres.
Palabras clave: eficacia de las vacunas; modelos animales; SARS-CoV-2; vacuna COVID-19.
Introduction:
In preclinical studies of the vaccine SOBERANA®02, it was essential to verify
the efficacy of the immune response against SARS-CoV-2 in animal models. In
a challenge assay, animals immunized with the vaccine antigen are exposed to
the virus in a controlled manner.
Objectives:
To evaluate the immunogenicity and protection of FR02 antigen against SARS-CoV-2
in the Syrian golden hamster model.
Methods:
Immunized hamsters were challenged with the viral variant D614G. Body weight,
respiratory signs, presence of viral RNA in nasopharynx and lung tissue were
monitored. Cultures were performed on Vero E6 cells and histopathological analysis
of the lungs. Unvaccinated infected animals and unvaccinated and uninfected
animals were used as controls.
Results:
Immunized hamsters showed no signs of disease; viral RNA concentrations in nasopharynx
and lungs were lower than unvaccinated infected hamsters, with reduced viral
load (6 logarithms) in the lungs on the day of peak viral replication, and undetectable
in 27.7%. A cytopathic effect was observed in 5.8% of viral cultures in the
lungs of immunized vs. non-immunized infected animals, in which it occurred
in 100%. Immunized hamsters had lower rates of acute lung damage and overall
histological damage.
Conclusions:
The immune response generated in immunized animals conferred protection against
infection, severe lung damage and the development of symptomatic disease in
hamsters.
Keywords: animal models; COVID-19 vaccine; SARS-CoV-2; vaccine efficacy.
Recibido: 15/03/2025
Aprobado:
29/04/2025
INTRODUCCIÓN
La infección por SARS-CoV-2, por su rápida propagación y letalidad en los primeros años de la pandemia de COVID-19, se consideró una emergencia de salud pública de importancia internacional.(1)
Los esfuerzos globales para controlarla, incluyeron el desarrollo de vacunas basadas en plataformas bien establecidas, para reducir los tiempos de desarrollo y generar confianza en su aplicación.(2) SOBERANA®02 se desarrolló sobre una plataforma tecnológica utilizada previamente en Cuba.(3) Es una vacuna conjugada a base de subunidades de proteínas, su antígeno FINLAY- FR-02 (FR02) es una proteína recombinante del dominio de unión al receptor (RBD) conjugada químicamente al toxoide tetánico (RBD6-TT) formulado en Al (OH)3.(4)
Para establecer un antígeno como candidato vacunal, los ensayos de reto en modelos animales son de gran importancia ya que permiten definir la respuesta inmunitaria del huésped a la infección y a la vacunación, por lo que se consideran un paso importante y crítico para proceder con los ensayos clínicos en humanos.(5)
Con el objetivo de probar la inmunogenicidad y eficacia protectora del antígeno FR02, se realizó un ensayo de reto viral con SARS-CoV-2 en el hámster sirio dorado.
MÉTODOS
Condiciones de seguridad
Los procedimientos con SARS-CoV-2 y animales infectados, se realizaron en una instalación de nivel de bioseguridad 3, del Centro de Investigaciones Científicas de la Defensa Civil, de Mayabeque, Cuba.
Virus
Se utilizó SARS-CoV-2, cepa D614G, aislamiento cubano, con secuencia genotípica referida en GISAID: hCov-19/Cuba/DC01/2020.
Modelo animal
Hámsteres sirio dorado (Mesocricetus auratus), 27 hembras (peso promedio de 116,5 g), organizados en 4 grupos, mantenidos en gabinetes ventilados, bajo régimen de luz y oscuridad por 12 horas, entre 20 y 24 0C, con acceso libre al agua y a los alimentos.
Grupos 1 y 2: 9 hámsteres por grupo, inmunizados con 5 µg y 3 µg de antígeno, respectivamente y retados.
Grupo 3: 6 hámsteres sin inmunizar, infectados con el virus (controles positivos).
Grupo 4: 3 hámsteres sin inmunizar y sin infectar (controles negativos).
Se realizó eutanasia después de 4 h de ayuno, en los días 3, 5 y 7 postinfección (PI), con sobredosis (80 mg/kg de peso animal) de tiopental sódico (Laboratorios AICA, La Habana, Cuba.). Se siguieron los estándares éticos para el trabajo con animales de laboratorio.(6,7,8,9)
Inmunización y reto
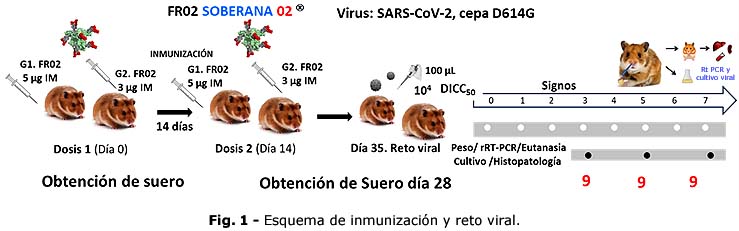
Se inmunizaron los animales con el antígeno FR02, por vía intramuscular (i.m.) con 2 dosis separadas por 14 días, con una concentración de 5 y 3 µg de antígeno, según grupo de estudio. El suero se obtuvo por punción del plexo retroorbitario a los 0 y 28 días (suero preinmune y 14 días después de la segunda inmunización) (Fig. 1).
Los animales inmunizados y los controles positivos, se inocularon por vía intranasal, con 100 µL del virus a una concentración de 104DICC50. Los hámsteres fueron pesados y anestesiados previamente con ketamina y xilacina (200 mg/kg y 10 mg/kg de peso respectivamente).
Observaciones clínicas
Se observó el estado físico de los animales diariamente, se evaluaron los signos respiratorios, peso corporal, actividad física y posturas. Los hámsteres se pesaron en el tiempo cero (T0), a los 3, 5 y 7 días PI, en una balanza digital (Sakura. GMBH, Gottingen. Alemania). Se estableció como línea base, el peso del T0 (día del reto)
Ensayo de neutralización viral
Se evaluó la capacidad neutralizante del suero de los hámsteres inmunizados, por un ensayo colorimétrico de neutralización del efecto citostático del SARS-CoV-2 en células Vero E6.(10) Las células se observaron en busca de signos de efecto citopático (ECP). Se visualizó la neutralización mediante la tinción con rojo neutro. Se consideró como título de neutralización la mayor dilución del suero evaluado con valor de densidad óptica superior al valor de corte. El valor de corte se calculó como el promedio de los valores de densidad óptica de los pocillos de control celular dividido por 2.
Toma de muestras, faríngeas y de pulmón
Se realizaron lavados de nasofaringe (NSF), en los días 3, 5, 7 PI, utilizando el medio MEM con antibiótico. Se tomaron alícuotas para la detección de ARN de SARS-CoV-2 por rRT-PCR y viabilidad viral por cultivo.
Los pulmones se examinaron macroscópicamente en busca de alteraciones anatomo-patológicas, cambios de coloración, congestión, focos hemorrágicos y edema. Se estudiaron además por histopatología, cultivo viral y rRT-PCR.
Procesamiento histológico
El análisis histopatológico se realizó según los procedimientos descritos por González S y otros.(11) Se determinó el área estimada de afectación del pulmón (AEAP) y se calculó el daño pulmonar agudo (DPA).(12) El daño pulmonar histológico global (DPG) se determinó mediante el método descrito por Li C y otros(13) en hámsteres infectados con SARS-CoV-2. Se calculó el porcentaje de inhibición del daño pulmonar en los grupos 1 y 2, considerando 100 % de daño la diferencia entre el grupo 3, control de infección, y el grupo 4, control negativo.(14)
Ensayo de reverso transcripción- reacción en cadena de la polimerasa en tiempo real (rRT- PCR) a partir de muestras de NSF y pulmones
La detección del virus se realizó utilizando el estuche RIDA ® GENE SARS-CoV-2 RUO (R-Biopharm AG).(15) Para la confirmación se emplearon los cebadores y sonda del estuche comercial Light Mix RdRP COVID-19 (TIB MOLBIOL, Roche), del gen de la polimerasa del SARS-CoV-2.(16) Se utilizó la plataforma para PCR, LightCycler 480 II (ROCHE).
Se determinaron los valores de ciclo umbral (Ct) (del inglés, cycle threshold ). El ensayo se consideró válido cuando los controles positivos mostraron curva sigmoidea, con valor de Ct 25,0 - 28,0 y los controles negativos sin señal de amplificación. Las muestras con curvas típicas de PCR (gen E Ct < 36 ciclos, gen RdRP < 40 ciclos) se consideraron positivas.(16)
Curva estándar de cuantificación de ARN viral
Para estimar la cantidad de ARN en las muestras se desarrolló una curva estándar para rRT-PCR(17) empleando el segundo estándar internacional de referencia de SARS-CoV-2 de la Organización Mundial de la Salud (OMS), con una concentración de 1 x 107 copias/mL de ARN viral (NIBSC código 22/52).(18) Se prepararon 6 puntos con concentraciones conocidas del analito y se determinó la relación lineal entre la concentración del analito y la señal, medida por el método de análisis (desde 1 x 107copias/mL hasta 1 x 102copias/mL), lo que posibilitó establecer la relación entre la cantidad de ARN y el ciclo de amplificación en la PCR.
Se realizó un análisis de regresión lineal, para determinar la ecuación de la recta que mejor se ajustara a los puntos experimentales. Se tuvieron en cuenta los parámetros de calidad de la curva estándar obtenida en el equipo de PCR , LightCycler 480 II (ROCHE).
Cultivo viral de las muestras de nasofaringe y pulmones
Las muestras se cultivaron durante 7 días en células Vero E6, incubadas a 37 °C en atmósfera de CO2al 5 %.(12) Las células se monitorearon diariamente para detectar cambios o aparición del ECP. Se confirmó la presencia de SARS-CoV-2 mediante rRT-PCR a partir del ARN viral de los sobrenadantes del cultivo.
Análisis estadístico
Los datos se analizaron mediante la aplicación informática GraphPad Prism (Versión 6.0, GraphPad Software, Inc. 2012).
Se utilizó la prueba ANOVA de dos vías y análisis post hoc mediante corrección de Bonferroni para comparaciones múltiples del peso y valores de Ct. Los títulos de anticuerpos se expresaron por animal individual y como la media ± DE (desviación estándar de la media) de cada grupo. Las diferencias significativas entre los grupos se determinaron mediante prueba de Kruskal-Wallis y la prueba de múltiples comparaciones de Dunn. Los valores probabilísticos (p) menores de 0,05 se consideraron significativos.
Aspectos bioéticos
Para realizar la investigación se tuvo en cuenta lo establecido en el Decreto Ley 31/2021 "De Bienestar Animal" del Consejo de Estado de la República de Cuba(6) y el Decreto 38/2021 "Reglamento del Decreto-Ley 31 de Bienestar Animal" del Consejo de Ministros de la República de Cuba.(7)
RESULTADOS
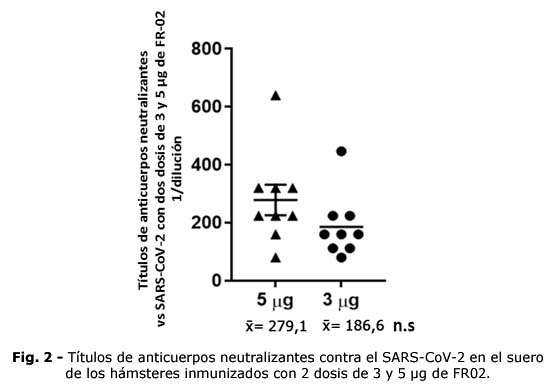
Los hámsteres inmunizados generaron anticuerpos que neutralizaron el virus SARS-CoV-2, tras 2 dosis de inmunización i.m. No hubo diferencias significativas entre los títulos de anticuerpos neutralizantes observados (Fig. 2).
Los animales inmunizados no presentaron lesiones en el lugar de la inyección, ni signos de enfermedad durante el reto, a excepción de estadios de somnolencia en el día 3 PI; a diferencia de los no inmunizados infectados, que manifestaron los signos descritos para el modelo, como postración, postura de espalda flexionada y respiración arrítmica, entre los días 3 y 5 PI. Los controles negativos no presentaron signos.
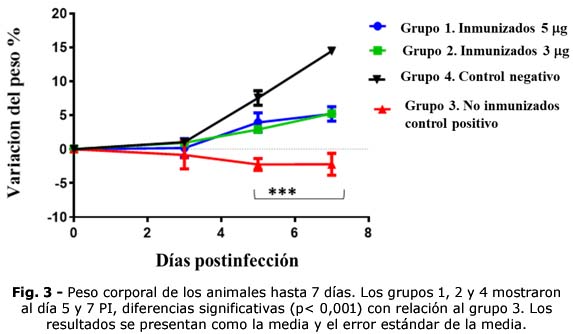
En los grupos 1 y 2 no disminuyó el peso corporal, ni hubo variaciones significativas entre ellos. A partir del día 5 PI tuvieron incrementos del peso (3,21 % y 4,73 %, respectivamente) con relación al T0 (Fig. 3).
Los animales del grupo 3, en los días 3, 5 y 7 PI disminuyeron el peso corporal, (promedios de variaciones de -2,79 %, -2,85 % y -2,21 %, respectivamente) y nunca lo recuperaron. Al comparar las diferencias del peso respecto al T0 se observaron diferencias significativas (p< 0,001) entre los animales de los grupos 1 y 2 con relación al grupo 3, en los días 5 PI y 7 PI. En los controles negativos se observaron mayores incrementos del peso corporal a partir del día 5 PI (6,8 %).
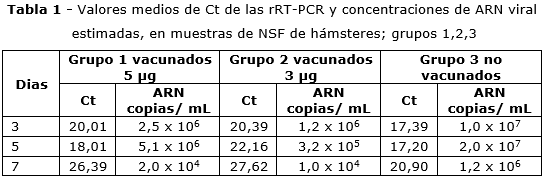
En la NSF de todos los hámsteres, a excepción de los controle negativos, se detectó ARN de SARS-CoV-2 en los días 3, 5 y 7 PI.
En los animales inmunizados, los mayores valores de carga viral (CV) fueron detectados en el día 5 PI (grupo 1) y en el día 3 PI (grupo 2). En ambos grupos, en el día 7 PI, se evidenció una disminución de la CV. Un hámster del grupo 2 mostró concentraciones < 102 copias/mL de ARN y un valor de Ct de 37,04 próximo al valor de corte (tabla 1).
En los animales no inmunizados infectados, en los días 3, 5 y 7 PI se observaron CV superiores a las apreciadas en los animales inmunizados. En el día 7 PI también ocurrió un corrimiento en las curvas de PCR, con reducción de la CV, pero permaneció mayor a la observada en inmunizados (tabla 1). Aunque hubo diferencias entre los valores de Ct y la concentración de ARN viral, en la NSF entre vacunados y no vacunados, estas no fueron significativas. En los controles negativos no se detectó ARN viral.
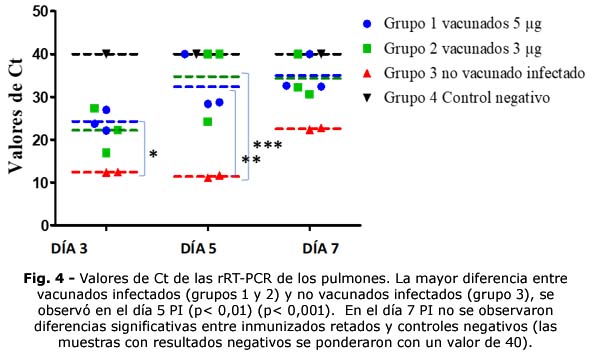
En los pulmones de los animales inmunizados (grupos 1 y 2) se observaron concentraciones de ARN viral inferiores a las apreciadas en NSF. En la figura 4 se puede observar cómo durante todo el proceso infeccioso, los valores de Ct de los animales inmunizados fueron mayores a los no vacunados infectados (grupo 3 ), lo que evidenció una CV menor.
En los grupos 1 y 2 en el día 3 PI se estimaron concentraciones de ARN viral de 8 x 104 y 3,2 x 105copias/mL de ARN, respectivamente, mientras que en los no vacunados infectados, las concentraciones fueron significativamente mayores (p< 0,05), con valores > 1 x 107copias/mL de ARN.
En el día 5 PI, se estimó la ocurrencia del pico de replicación viral en el grupo 3, sugerido por el valor medio más bajo de Ct observado (11,40) y concentración de ARN viral estimada> 1 x 107 copias/mL. En ese tiempo, en los hámsteres inmunizados (grupos 1 y 2) se observaron concentraciones menores de CV (3,1 x 10 2 y < 1 x 10 2 copias/mL de ARN viral, respectivamente), con diferencias significativas en los valores de Ct (p< 0,01 y p< 0,001, respectivamente). Un animal del grupo 1 y en dos del grupo 2, no se detectó material genómico del virus en los pulmones (Fig. 4). Entre los grupos inmunizados 1 y 2 no hubo diferencias significativas.
En el día 7 PI los animales inmunizados mostraron valores de Ct representativos de un estadio próximo a negativizar el proceso infeccioso, con CV baja (< 102 copias/mL de ARN ) y no tuvieron diferencias significativas respecto al grupo control negativo. Un hámster de cada grupo vacunado no evidenció presencia de ARN viral (Fig. 4), mientras que en el grupo de no inmunizados infectados, persistieron valores elevados de CV (3,2 x 105 copias/mL de ARN). El grupo 4 se mantuvo negativo todo el ensayo.
En los cultivos de NSF de los animales de los grupos 1, 2 y 3 se observó ECP sobre las células Vero E6 (excepto un animal del grupo 2 en el día 7 PI).
En los cultivos de pulmón del grupo 1 no se observó ECP en los días 3,5,7 PI; igual ocurrió en los animales del grupo 2, con excepción de una muestra que mostró ECP y otra, en la cual no se pudo determinar, ambas en el día 5 PI. En el cultivo viral de los pulmones de los animales inmunizados (grupos 1 y 2) solo se observó ECP en el 5,8 % de las muestras de pulmón, a diferencia del grupo 3 en el cual se observó en el 100 % de las muestras. Las muestras de NSF y pulmón del grupo 4, resultaron negativas.
Macroscópicamente los pulmones de los hámsteres inmunizados, presentaron pequeñas zonas de enrojecimiento y punteado hemorrágico. En todos los casos, se apreciaron menos lesiones con relación al grupo 3 no inmunizado; se visualizaron zonas necróticas, coágulos y edemas. En los pulmones de los hámsteres del grupo 4 no se observaron lesiones.
En la histopatología se observó menor afectación en los pulmones de los hámsteres inmunizados, respecto a los no inmunizados infectados, que presentaron lesiones en aumento desde el día 3 hasta el día 7 PI, caracterizadas por engrosamiento grave de las paredes alveolares, congestión de los capilares alveolares con infiltrado inflamatorio. En el interior de los alveolos se apreció material proteináceo, hemorragias y células multinucleadas, neumocitos de tipo I y tipo II. Se evidenció atelectasia, enfisema focal, afectación del 80 % del parénquima y neumonía intersticial aguda grave extensa .
El grupo 1 tuvo menores AEAP que el grupo 2, lo que se correspondió con mayores porcentajes de inhibición del daño pulmonar. En el día 5 PI en el grupo 3 se observó un AEAP del 60 %, mientras que en el grupo 1 fue de 27,5 %, con un porcentaje de inhibición del 100 %. De igual manera, en el grupo 3 se observaron los mayores puntajes de DPA, (0,11 en el día 5 PI), mientras que en los grupos 1 y 2 fue menor (0,07 y 0,09 respectivamente). El mayor porcentaje de inhibición del DPA respecto al grupo 3, se observó en el grupo 1. En el grupo 3 se observaron los mayores índices de DPG en los tiempos evaluados.Los mejores resultados de inhibición del DPG se apreciaron en el grupo 1 en los días 5 y 7 PI.
DISCUSIÓN
El grado de afinidad de la unión entre el RBD de la proteína S del virus y el receptor ACE2, determina la eficiencia de ingreso del virus en la célula hospedera,(20) por lo que la capacidad de los anticuerpos de neutralizar esta interacción es un criterio de funcionalidad.(21) Las dos concentraciones del antígeno FR02 utilizadas para inmunizar los animales, fueron capaces de generar títulos de anticuerpos neutralizantes contra el virus y proteger a los animales de los grupos 1 y 2 de los signos característicos de la enfermedad.
Por el contrario, los animales no inmunizados infectados, además de enfermarse redujeron en un 2 % su peso, lo que biológicamente resulta significativo para un animal pequeño, en un corto período. La pérdida del peso fue inferior a la observada en estudios realizados previamente,(12) lo que puede deberse al sexo de los hámsteres; en las hembras se describen menores pérdidas que en los machos.(22,23)
En la NSF, de los animales inmunizados con 5 µg y 3 µg de FR02, aunque ocurrió poca reducción de la concentración de ARN viral, respecto a los controles positivos. La disminución de 1 logaritmo, en el contexto de la cuantificación de material genético en estudios virológicos es considerada alta, además de que en el escenario de la COVID-19, la reducción de CV en NSF resulta importante a la hora de disminuir el riesgo de contagio.(24) La disminución de la CV a los 7 días PI, puede interpretarse como el inicio de la recuperación; resultados similares a los reportados por otros autores.(19,25,26)
Estas diferencias de CV en NSF entre vacunados y no vacunados se ha observado en estudios similares con animales inmunizados con antígenos de subunidades proteicas y animales no inmunizados.(27)
Resultó notable, la reducción de la CV en los pulmones de los animales vacunados desde el día 3 PI, en el orden de 3 a 4 logaritmos respecto a los controles positivos, lo que fue más significativo en el día 5 PI, fecha en que ocurrió el pico de replicación viral en los animales no vacunados. Las concentraciones de ARN viral observadas en los hámsteres inmunizados evidenciaron un aclaramiento viral en el orden de 6 logaritmos, incluso hubo 3 hámsteres, en que no se detectó ARN viral en los pulmones. En el 100 % de los hámsteres sin inmunizar infectados se detectó material genómico del virus.
Estos hallazgos indican el nivel de protección conferida por la inmunización y resultan importantes para prevenir la enfermedad grave y la muerte, según lo observado en pacientes con la COVID-19, que al ingresar con valores altos de CV, ocurría mayor mortalidad intrahospitalaria en comparación con quienes presentaban una CV baja.(28)
Estudios similares con vacunas contra el SARS-CoV-2, realizados en hámsteres, no observaron diferencias significativas entre vacunados y no vacunados en NSF, pero en los pulmones de todos los vacunados comprobaron una significativa disminución de las cargas virales.(29,30,31)
Los resultados de las rRT-PCR de NSF de todos los animales se correspondió con lo observado en los cultivos, confirmando la presencia de virus replicativos; sin embargo en los cultivos de pulmones de los grupos 1 y 2 no ocurrió igual, debido a la dificultad del cultivo virológico a partir de muestras con baja CV, en contraste con la alta sensibilidad de la PCR para detectar cantidades ínfimas de ARN viral.(32,33,34)
Los resultados positivos de los cultivos y de los ensayos de rRT-PCR en las muestras de pulmones del grupo 3, se correlacionan con los análisis histopatológicos. En los animales no inmunizados infectados en los días en los cuales se observaron las mayores lesiones según el puntaje DPA, fueron detectadas cargas virales altas en los pulmones. Esto coincide con lo publicado por otros autores, que reportaron el pico de patología y consolidación pulmonar en el día 7 postinfección,(19,35,36) o días después de ocurrir el pico replicativo viral.(34,37)
La ausencia de signos característicos de la enfermedad y de virus infectivos en los cultivos celulares, así como de menores lesiones histopatológicas en los animales inmunizados, principalmente los del grupo 1, respecto al grupo control de infección, habla a favor de la protección conferida por el antígeno de SOBERANA®02, incluso cuando la vacuna se administró en un corto período antes de la exposición al virus.
Las inmunizaciones ensayadas resultaron eficaces y generaron respuestas de anticuerpos que neutralizaron el virus SARS-CoV-2, redujeron la CV, el daño tisular y la inflamación en los pulmones. Se demostró la capacidad del candidato vacunal de prevenir estadios graves de la enfermedad y la enfermedad sintomática en los hámsteres, pues la respuesta inmune generada en los animales inmunizados confirió protección contra la infección, el daño pulmonar grave y el desarrollo de la enfermedad sintomática en los hámsteres.
Agradecimientos
Los autores agradecen al zootecnista Alex Quintero Pérez del IFV la encomiable labor desempeñada en la atención y manejo de los animales de laboratorio, así como al colectivo de ingenieros y técnicos del CICDC que garantizaron las condiciones de confort y de contención física de la instalación con nivel de bioseguridad 3.
REFERENCIAS BIBLIOGRÁFICAS
1. Organización Mundial de la Salud. Declaración sobre la segunda reunión del Comité de emergencias del reglamento Sanitario Internacional acerca del brote del nuevo coronavirus (2019-nCoV) [Internet]. Ginebra: OMS; 2020. [acceso: 30/01/2020]. Disponible en: https://www.who.int/es/news/item/30-01-2020-statement-on-the-second-meeting-of-the-international-health-regulations-(2005)-emergency-committee-regarding-the-outbreak-of-novel-coronavirus-(2019-ncov)
2. Nagy A, Alhatlani B. An overview of current COVID-19 vaccine platforms [Internet]. Comput Struct Biotechnol J. 19:2508-17. 2021. DOI: 10.1016/j.csbj.2021.04.061
3. Carlson R, Lutmer H, Beall M, Bolt B, Felton C, Germann L, et al. Precision Vaccinations. Cuba Covid-19 Vaccine. [Internet]. Vax-Before-Travel.2022[acceso: 06/05/2022]. Disponible en: https://www.vax-before-travel.com/vaccines/cuba-covid-19-vaccine
4. Valdes-Balbin Y, Santana-Medero D, Quintero L, Fernández S, Rodriguez L, Sanchez Ramirez B, et al. SARS-CoV-2 RBD-tetanus toxoid conjugate vaccine induces a strong neutralizing immunity in preclinical studies [Internet]. ACS Chem Biol. 2021;16:1223-33. DOI: 10.1101/2021.02.08.430146
5. Griffin JFT. A strategic approach to vaccine development: animal models, monitoring vaccine efficacy, formulation and delivery [Internet]. Adv. Drug Del. Rev. 2002; 54: 851-861. DOI: 10.1016/S0169-409X(02)00072-8
6. Consejo de Estado República de Cuba. Decreto-Ley 31/2021 "De Bienestar Animal". Gaceta Oficial No. 25 Extraordinaria de 10 abril de 2021 [Internet]. La Habana: Ministerio de Justicia; 2021. [acceso: 06/05/202]. Disponible en: https://www.mined.gob.cu/wp-content/uploads/2022/01/goc-2021-ex25-.pdf
7. Consejo de Estado República de Cuba. Decreto 38/2021 "Reglamento del Decreto-Ley de Bienestar Animal". Gaceta Oficial No. 25 Extraordinaria de 10 abril de 2021 [Internet]. La Habana: Ministerio de Justicia; 2021. [acceso: 06/05/202]. Disponible en: https://www.mined.gob.cu/wp-content/uploads/2022/01/goc-2021-ex25-.pdf
8. Committee for the Update of the Guide for the Care and Use of Laboratory Animals. Institute for Laboratory Animal Research. Division on Earth and Life Studies. National Research Council of the National Academies. The National Academies Press. Guide for the Care and Use of Laboratory Animals [Internet]. Eighth Edition. Washington, D.C.: National Academies of Sciences, Engineering, and Medicine; 2011. DOI: 10.17226/12910
9. American Veterinary Medical Association. AVMA Guidelines for the Euthanasia of Animals: 2020 Edition. Schaumburg: American Veterinary Medical Association; 2020. [acceso: 06/05/2022]. Disponible en: https://olaw.nih.gov/policies-laws/avma-guidelines-2020.htm
10. Manenti A, Maggetti M, Casa E, Martinuzzi D, Torelli A, Trombetta CM, et al. Evaluation of SARS-CoV-2 neutralizing antibodies using of a CPE-based colorimetric live virus micro-neutralization assay in human serum samples [Internet]. J Med Virol. 2020; 1-9. DOI: 10.1002/jmv.25986
11. González-Sosa N, Oliva-Hernández R, Blanco-de Armas M, Infante-Bouzac J, Enríquez-Puertas J, Rodríguez-Salgueiro S, et al. Infectividad del coronavirus tipo 2 del síndrome respiratorio agudo severo en hámster Sirio Dorado como modelo para ensayos de inmunogenicidad y eficacia de biomoléculas y candidatos vacunales [Internet]. Vaccimonitor. 2024 [acceso: 17/12/2024];33:e153324. Disponible en: https://vaccimonitor.finlay.edu.cu/index.php/vaccimonitor/article-/view/9218
12. Matute-Bello G, Downey G, Moore BB, Groshong SD, Matthay MA, Slutsky AS, et al. Acute Lung Injury in Animals Study Group. An official American Thoracic Society workshop report: features and measurements of experimental acute lung injury in animals [Internet]. Am J Respir Cell Mol Biol. 2011; 44(5):725-38. DOI: 10.1165/rcmb.2009-0210ST
13. Li C, Chen YX, Liu FF, Lee AC, Zhao Y, Ye ZH, et al. Absence of Vaccine-enhanced Disease with Unexpected Positive Protection Against severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) by Inactivated Vaccine Given Within 3 Days of Virus Challenge in Syrian Hamster Model [Internet]. Clin Infect Dis. 2021;73(3): e719-e734. DOI: 10.1093/cid/ciab083
14. Molina-Cuevas V, Ravelo-Calzado Y, Zamora-Rodríguez Z, Noa-Puig M, Valle-Clara M, Pérez-Guerra, Y, et al. Efectos en ratas de los alcoholes de cera de abejas (D-002) sobre la colitis ulcerativa inducida por sulfato de dextrano y etanol [Internet]. Revista Peruana de Medicina Experimental y Salud Pública. 2017; 34(2):176-182. DOI: 10.17843/rpmesp.2017.342.2369
15. R-Biopharm AG. RIDA®GENE SARS-CoV-2 REF. PG6820. Alemania, Darmstadt; 2020. [acceso: 17/12/2024]. Disponible en: https://clinical.r-biopharm.com/wp-content/uploads/2020/06/pg6820_ridagene_sars-cov-2_2020-10-28_en.pdf
16. Nalla AK, Casto AM, Huang M-LW, Perchetti GA, Sampoleo R, Shrestha L, et al. Comparative performance of SARS-CoV-2 detection assays using seven different primer- probe sets and one assay kit [Internet]. J Clin Microbiol. 2020; 58: e00557-20. DOI: 10.1128/JCM.00557-20
17. Sahoo M, Huang C, Sibai M, Solis D, Pinsky B. Harmonization of SARS-CoV-2 reverse transcription quantitative PCR tests to the first WHO international standard for SARS-CoV-2 RNA [Internet]. Journal of Clinical Virology. 2022;154. DOI: 10.1016/j.jcv.2022.105242.105242
18. Bentley EM, Le Duff Y, Ham C, Cherry C, Mattiuzzo G, Fryer J, et al. Collaborative Study for the Establishment of the Second WHO International Standard for SARS-CoV-2 RNA [Internet]. Geneva: World Health Organization Expert Committee on Biological Standardization. WHO/BS/2023.2459; 2023. [acceso: 17/12/2024]. Disponible en: https://www.who.int/publications/m/item/who-bs-2023.2459
19. Chan JF, Zhang AJ, Yuan S, Poon VK, Chan CC, Lee AC, et al. Simulation of the Clinical and Pathological Manifestations of Coronavirus Disease 2019 (COVID-19) in a Golden Syrian Hamster Model: Implications for Disease Pathogenesis and Transmissibility [Internet]. Clin Infect Dis. 2020;71(9):2428-2446. DOI: 10.1093/cid/ciaa325
20. Wan Y, Shang J, Graham R, Baric RS, Li F. Receptor Recognition by the Novel Coronavirus from Wuhan: an Analysis Based on Decade-Long Structural Studies of SARS Coronavirus [Internet]. J Virol. 2020; 94(7): e00127-20. DOI: 10.1128/JVI.00127-20
21. Tan CW, Chia WN, Qin X, Liu P, Chen MI, Tiu C, et al. A SARS-CoV-2 surrogate virus neutralization test based on antibody-mediated blockage of ACE2-spike protein-protein interaction [Internet]. Nat Biotechnol. 2020; 38(9):1073-8. DOI: 10.1038/s41587-020-0631-z
22. Griffin BD, Warner BM, Chan M, Valcourt E, Tailor N, Banadyga L, et al. Host parameters and mode of infection influence outcome in SARS-CoV-2-infected hamsters [Internet]. iScience. 2021;24(12):103530. DOI: 10.1016/j.isci.2021.103530
23. Yuan L, Zhu H, Zhou M, Ma J, Chen R, Chen Y, et al. . Gender associates with both susceptibility to infection and pathogenesis of SARS-CoV-2 in Syrian hamster [Internet]. Signal Transduct Target Ther. 2021;6(1):136. DOI: 10.1038/s41392-021-00552-0
24. Cuenca-Pardo J, Ramos-Gallardo G, Vélez-Benítez E, Álvarez-Díaz Carlos de J, Bucio-Duarte J, Iribarren-Moreno R, et al. La importancia de reducir la carga viral para disminuir el riesgo de contagio por COVID-19 [Internet]. Cir Plast. 2020;30(2):78-93. DOI:10.35366/97674
25. Sia SF, Yan LM, Chin AWH, Fung K, Choy KT, Wong AYL, et al. Pathogenesis and transmission of SARS-CoV-2 in golden hamsters. Nature. 2020;583(7818): 834-838. DOI: 10.1038/s41586-020-2342-5
26. Lee AC, Zhang AJ, Chan JF, Li C, Fan Z, Liu F, et al. Oral SARS-CoV-2 Inoculation Establishes Subclinical Respiratory Infection with Virus Shedding in Golden Syrian Hamsters [Internet]. Cell Rep Med. 2020;1(7):100121. DOI: 10.1016/j.xcrm
27. Lin YJ, Lin MY, Chuang YS, Liu LT, Kuo TY, Chen C, et al Protection of hamsters challenged with SARS-CoV-2 after two doses of MVC-COV1901 vaccine followed by a single intranasal booster with nanoemulsion adjuvanted S-2P vaccine. Sci Rep. 2022;12(1):11369. DOI: 10.1038/s41598-022-15238-y
28. Vega-de LoPresti Y, Montilva-Gutiérrez E, Valenzuela-Vegas A, Salazar-Pérez KE, Paredes-Manodanda AS, Alvarado-García AJ, et al. Upper respiratory tract viral load quantification in COVID-19 patients at hospital admission and its association with disease severity [Internet]. Gac Med Mex. 2022;158(5):320-326. DOI: 10.24875/GMM.M22000700
29. O'Donnell KL, Clancy CS, Griffin AJ, Shifflett K, Gourdine T, Thomas T, et al. Optimization of Single-Dose VSV-Based COVID-19 Vaccination in Hamsters. Frontiers in Immunology. 2022; 12:788235 DOI: 10.3389/fimmu.2021.788235
30. Yahalom-Ronen Y, Tamir H, Melamed S, Politi B, Shifman O, Achdout H, et al. A single dose of recombinant VSV-?G-spike vaccine provides protection against SARS-CoV-2 challenge [Internet]. Nat Commun. 2020;11(1):6402. DOI: 10.1038/s41467-020-20228-7
31. Wussow F, Kha M, Faircloth K, Nguyen VH, Iniguez A, Martinez J, et al. COH04S1 and beta sequence-modified vaccine protect hamsters from SARS-CoV-2 variants [Internet]. iScience. 2022;25(6):104457. DOI: 10.1016/j.isci.2022.104457
32. Jefferson T, Spencer EA, Brassey J, Heneghan C. Viral Cultures for Coronavirus Disease 2019 Infectivity Assessment: A Systematic Review[Internet]. Clin Infect Dis. 2021;73(11):e3884-e3899. DOI: 10.1093/cid/ciaa1764
33. Jiang RD, Liu MQ, Chen Y, Shan C, Zhou YW, Shen XR, et al. Pathogenesis of SARS-CoV-2 in Transgenic Mice Expressing Human Angiotensin-Converting Enzyme 2 [Internet]. Cell. 2020;182(1):50-58. DOI: 10.1016/j.cell.2020.05.027
34. Chandrashekar A, Liu J, Martinot AJ, McMahan K, Mercado NB, Peter L, et al. SARS-CoV-2 infection protects against rechallenge in rhesus macaques [Internet]. Science. 2020;369(6505):812-17. DOI: 10.1126/science.abc4776
35. Muñoz-Fontela C, Dowling WE, Funnell SGP, Gsell PS, Riveros-Balta AX, Albrecht RA, et al. Animal models for COVID-19 [Internet]. Nature. 2020;586(7830):509-15. DOI: 10.1038/s41586-020-2787-6
36. Imai M, Iwatsuki-Horimoto K, Hatta M, Loeber S, Halfmann PJ, Nakajima N, et al. Syrian hamsters as a small animal model for SARS-CoV-2 infection and countermeasure development [Internet]. Proc Natl Acad Sci USA. 2020;117(28):16587-95. DOI: 10.1073/pnas.2009799117
37. Rosenke K, Meade-White K, Letko M, Clancy C, Hansen F, Liu Y, et al. Defining the Syrian hamster as a highly susceptible preclinical model for SARS-CoV-2 infection [Internet]. Emerg Microbes Infect. 2020;9(1):2673-84. DOI:10.1080/22221751.2020.1858177
Conflictos de interés
Los autores declaran que no existen conflictos de interés en relación con el trabajo.
Contribuciones de los autores
Conceptualizacion: Nibaldo Luis González Sosa, Reynaldo Oliva, Madeline
Blanco de Armas.
Curación
de datos: Nibaldo Luis González Sosa, Reynaldo OlivaHernández, Madeline Blanco,Mildrey
Fariñas Medina,Juan Francisco Infante Bouzac,Yanet Climent Ruiz,Sonsire Fernández
Castillo, Liuber Yans Machado Zaldívar, Enrique Noa Romero.
Análisis
formal: Nibaldo Luis González Sosa, Reynaldo OlivaHernández, Madeline Blanco
de Armas,Mildrey Fariñas Medina,Juan Francisco Infante Bouzac, Yanet Climent
Ruiz,Sonsire Fernández Castillo.
Investigación:
Nibaldo Luis González Sosa, Reynaldo OlivaHernández, Madeline Blanco,Mildrey
Fariñas Medina,Juan Francisco Infante Bouzac,Yanet Climent Ruiz,Sonsire Fernández
Castillo,Liuber Yans Machado Zaldívar, Enrique Noa Romero, Darcy Núñez Martínez,
Sandra Rodríguez Salgueiro, Yisabel Aranguren Mazorra.
Metodología:
Nibaldo Luis González Sosa, Reynaldo Oliva, Madeline Blanco de Armas,
Juan Francisco Infante Bouzac.
Administración
del proyecto: Dagmar García Rivera, Nibaldo Luis González Sosa, Reynaldo
Oliva.
Supervisón:
Dagmar García Rivera, Mireida Rodríguez Acosta, Otto Cruz Sui, Marta
Dubed Echevarría.
Redacción - Redacción-borrador original: Nibaldo Luis González Sosa, Madeline
Blanco de Armas.
Redacción-revisión
y edición: Nibaldo Luis González Sosa, Madeline Blanco de Armas, Dagmar
García Rivera,Liuber Yans Machado Zaldívar , Mireida Rodríguez Acosta,
Marta Dubed Echevarría.
Declaración de disponibilidad de datos
Los datos suplementarios de la investigación no incluidos en el artículo están disponibles para su consulta, previo contacto con el primer autor al correo: nibaluis@gmail.com
Archivo complementario: Tablas complementarias; formato PDF. Disponible en: https://revmedmilitar.sld.cu/index.php/mil/libraryFiles/downloadPublic/47