Artículo de Investigación
Efecto de la variedad de maíz transgénica H-Ame15 en la salud del suelo e implicación en la seguridad alimentaria
Effect of the transgenic corn variety H-Ame15 on soil health and implications for food security
Odette Beiro Castro1* https://orcid.org/0000-0002-2403-5799
Yordanka Dominguez Linares1 https://orcid.org/0000-0001-6199-8757
Ana Cristina Noa Rodríguez1 https://orcid.org/0000-0003-2857-1385
Pilar Tellez-Rodriguez2 https://orcid.org/0000-0001-7572-3498
Licette Leon Barreras2 https://orcid.org/0009-0009-1673-5670
Aymee Ferrer Colás1 https://orcid.org/0009-0004-2140-698X
Mario Pablo Estrada García2 https://orcid.org/0000-0002-9776-719X
1Centro Nacional de Toxicología. La Habana, Cuba.
2Centro de Ingeniería Genética y Biotecnología. La Habana, Cuba.
*Autor para la correspondencia. Correo electrónico: odettebeiro1968@gmail.com
RESUMEN
Introducción: Evaluar la fauna del suelo es clave para comprender el impacto del maíz híbrido H-Ame15 en la salud edáfica, directamente relacionada con la calidad y seguridad de los alimentos. El deterioro de la biodiversidad altera los ciclos biogeoquímicos, compromete la nutrición de los cultivos y, puede afectar la salud humana, siendo fundamental para garantizar alimentos seguros.
Objetivo: Analizar el efecto del maíz híbrido H-Ame15 y las proteínas insecticidas Cry1Fa y Vip3A en bioindicadores del suelo.
Métodos: Ensayo desarrollado según la guía 850.3200. Grupos experimentales: Control (glucosa), proteínas inactivas y activas, residuos vegetales del híbrido y su contraparte no transgénica y dos con lombrices de tierra (Eisenia andrei). Se evaluaron los efectos en la comunidad microbiana mediante la cuantificación de CO2 en los días 5, 28, 35, 42 y 63 (final del ensayo). En las lombrices se analizó la mortalidad y signos de toxicidad, con frecuencia semanal.
Resultados: La emisión de CO2 no se modificó con las proteínas. El maíz híbrido mostró menores niveles de respiración que el convencional (-0,96 mg/g en el día 63). La presencia de lombrices favoreció la función respiratoria: Valores de 20,22 y 18,54 mg/g para los maíces, convencional e hibrido, respectivamente. En las lombrices no se evidenció toxicidad.
Conclusión: El maíz híbrido H-Ame15 y las proteínas que expresa no provocaron daños relevantes en los indicadores de la salud edáfica, lo cual sugiere que no existirían riesgos indirectos para la salud humana asociados a su cultivo.
Palabras clave: maíz; plantas modificadas genéticamente; respiración microbiana; lombrices de tierra; pruebas de toxicidad.
ABSTRACT
Introduction: Evaluating soil fauna is key to understanding the impact of hybrid corn H-Ame15 on soil health, which is directly related to food quality and safety. Biodiversity degradation alters biogeochemical cycles, compromises crop nutrition, and can affect human health, making it essential to ensure safe food.
Objective: To analyze the effect of hybrid corn H-Ame15 and the insecticidal proteins Cry1Fa and Vip3A on soil bioindicators.
Methods: The assay was developed according to guideline 850.3200. Experimental groups: Control (glucose), inactive and active proteins, plant residues from the hybrid and its non-transgenic counterpart, and two groups with earthworms (Eisenia andrei). The effects on the microbial community were assessed by quantifying CO2 on days 5, 28, 35, 42, and 63 (end of the assay). Earthworms were analyzed weekly for mortality and signs of toxicity.
Results: CO2 emissions were not modified by proteins. Hybrid maize showed lower respiration levels than conventional maize (-0.96 mg/g on day 63). The presence of earthworms favored respiratory function: Values of 20.22 and 18.54 mg/g for conventional and hybrid maize, respectively. No toxicity was evident in the earthworms.
Conclusion: Hybrid maize H-Ame15 and the proteins it expresses did not cause significant damage to soil health indicators, suggesting that there are no indirect risks to human health associated with its cultivation.
Keywords: corn; genetically modified plants; microbial respiration; earthworms; toxicity tests.
Recibido: 06/06/2025
Aprobado: 01/09/2025
INTRODUCCIÓN
La aparición y el resurgimiento de enfermedades, la creciente preocupación de la población por la inocuidad de los alimentos y el temor a posibles pandemias, han contribuido a la toma de conciencia de la estrecha interacción entre la salud humana, la sanidad animal y el medio ambiente. El desarrollo del concepto de “Una sola salud” aborda abordar los problemas sanitarios de forma integral, con una perspectiva interdisciplinaria que se dirige a minimizar los daños y maximizar los beneficios.(1) Este enfoque pretende desarrollar estrategias más eficientes y eficaces para solucionar problemas de salud, en la interrelación entre los seres humanos, los animales y el medio ambiente.(2)
La agricultura enfrenta el desafío de asegurar un cantidad adecuada de alimentos y preservar el equilibrio ambiental ante el aumento de la población mundial.(3) El uso de la biotecnología en la producción de alimentos es una de las esperanzas para abordar este desafío; se destacan los cultivos transgénicos, por sus fortalezas, pero sin desestimar los riesgos para la salud humana y ambiental.(4,5,6)
La estimación del riesgo ambiental (ERA) de un cultivo transgénico se basa en el análisis de factores como el receptor, la modificación genética, el organismo modificado genéticamente, la liberación o uso previsto, el posible entorno receptor y la interacción entre estos.(1) En este sentido, la evaluación de la fauna del suelo es clave para comprender el impacto de estos cultivos en la salud humana. Un deterioro en la biodiversidad aumenta los riesgos de contaminantes, afecta la calidad nutricional de los alimentos y pone en peligro la salud pública. Asegurar que el suelo mantenga su fauna saludable es fundamental para proteger la salud y garantizar alimentos seguros.(6)
Las comunidades microbianas y la lombriz de tierra, por su rol en los procesos y funciones que tienen lugar en el suelo, se consideran, entre otros aspectos, para la afectación en bioindicadores de la salud de este.(2,7)
En Cuba, el Centro de Ingeniería Genética y Biotecnología (CIGB) es la institución líder en la obtención de cultivos transgénicos, como el maíz híbrido H-Ame15, que se desarrolló para el control del lepidóptero Spodoptera frugiperda, conocido como palomilla del maíz. La resistencia de este híbrido a dicho insecto-plaga, se obtiene a partir de la expresión de las proteínas transgénicas, Cry1Fa y Vip3A provenientes de Bacillus thuringiensis (Bt) y aportadas por los eventos transgénicos TC1507 y MIR162. La combinación de ambas toxinas en estas plantas constituye además, una estrategia para el manejo de la posible aparición de insecto-resistencia.(8)
En el caso del híbrido, al cumplir los requisitos mínimos identificados, para extrapolar los datos de inocuidad obtenidos en la ERA de las líneas que contienen los eventos simples TC1507 y MIR162(9,10,11) se extrapolaron los resultados, pero es preciso completar el vacío en cuanto al impacto en las comunidades microbianas.(12)
El objetivo de esta investigación es analizar en el laboratorio el efecto del maíz híbrido H-Ame15 en bioindicadores de la salud del suelo.
MÉTODOS
Se realizó un ensayo ecotoxicológico en comunidades microbianas del suelo y en la lombriz de tierra. Se llevó a cabo en la Subdirección de Evaluaciones Toxicológicas y Medio Ambiente (SETMA), del Centro Nacional de Toxicología.
El ensayo se desarrolló por la guía 850.3200 de la Oficina de Seguridad Química y Prevención de la Contaminación (OCSPP, del inglés). El sistema fue estático, en condiciones de oscuridad.
El estudio tuvo una duración de 63 días.
Materiales
Maíz: El híbrido H-Ame15 se obtuvo a partir del cruce entre la línea L-Moltó, que contiene los eventos transgénicos TC1507 y MIR162 con la línea convencional cubana (CT9), para garantizar caracteres de resistencia, tolerancia y adaptación a las condiciones del clima. Se utilizaron, por frasco, 40 g de muestras de tejido senescente de maíz convencional e híbrido.
Proteínas insecticidas: Cry1Fa y Vip3Aa20 activas e inactivas; estas últimas, luego de someterlas a 65 °C por 10 minutos. Las concentraciones de las proteínas en el sustrato, correspondieron a niveles 10 veces por encima de la expresión del evento apilado.
Las muestras de vegetales y proteínas provinieron las suministró el CIGB.
Sustrato: Se empleó el humus de lombriz, para garantizar la carga microbiana.
Biomodelos: Lombrices de tierra de la especie Eisenia andrei, sanas, sexualmente maduras y mayores de 2 meses de edad. Se usaron 60 organismos, distribuidos en 2 grupos de 30 cada uno, a razón de 10 organismos por réplica, y 3 réplicas por grupo. La lombriz de tierra y el sustrato procedieron del área de cría y mantenimiento de biomodelos de la SETMA.
Variables
pH del soporte de investigación. Se midió al inicio y final del ensayo, para ello se pesó 1 g de sustrato en una balanza Sartorius, se enrazaron a 99 mL de agua destilada, se agitaron y dejaron reposar; se determinó la concentración de iones hidrógeno presente en las disoluciones mediante el empleo del pHmetro marca InoLab.
Emisión de CO2 de la comunidad microbiana. Se midió el primer día del experimento en el grupo control al que se le suministró glucosa, a partir del quinto día se midió en los demás grupos; los expuestos a las proteínas activas e inactivas hasta el día 28, en el resto además los días 35, 42 y 63.
Toxicidad en la lombriz de tierra (se analizaron semanalmente):
Mortalidad.
Alteraciones fisiológicas.
Alteraciones conductuales.
Variación del peso (del inicio vs. final del estudio).(13)
Procedimientos
La exposición fue desde el inicio, mediante el contacto e ingestión del sustrato. Para estimular la función respiratoria de la comunidad microbiana se añadió a todos los grupos glucosa. Para evaluar el efecto sobre la lombriz de tierra, así como en el proceso de descomposición. Para ello se usaron diferentes grupos experimentales cuyos tratamientos y concentraciones de las proteínas se muestran en la tabla1.
Procesamiento estadístico
Se empleó el paquete de análisis SPSS versión 26. Luego de probarla normalidad y homocedasticidad de la variable emisión de CO2 de la comunidad microbiana se le aplicó la prueba de comparación múltiple de Duncan, para analizar las medias de los grupos e identificar las diferencias.(14)
RESULTADOS
En todo momento los valores registrados de pH (7.92 - 8.5) fueron adecuados para el desarrollo de los microorganismos y las lombrices en ensayo.
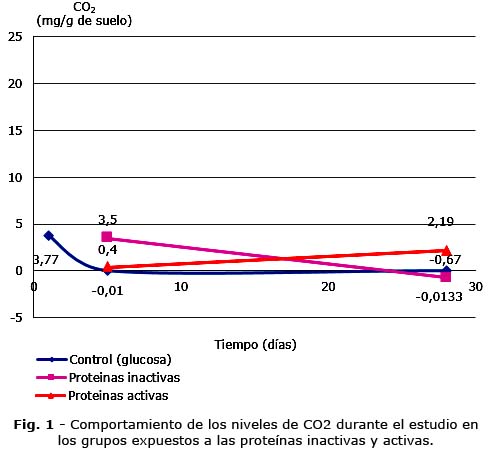
Los resultados de la cuantificación de CO2 en los grupos expuestos a las proteínas activas e inactivas se representan en la figura 1.
En el día cinco, al aplicar el procesamiento estadístico existieron diferencias significativas entre el grupo con las proteínas inactivas en contraste con los grupos control (glucosa) y el expuesto a las proteínas activas (p< 0,5), diferencias relacionadas con el hecho de que las proteínas inactivas estimularon la respiración en la comunidad microbiana. En el día 28 del estudio los análisis no revelaron diferencias estadísticamente significativas entre las medias de los niveles de CO2, lo que sugiere que la respiración fue similar entre ellos (p≥ 0,5).
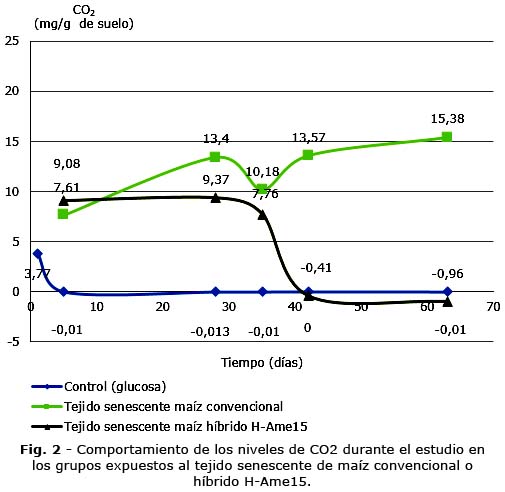
En la figura 2 se constata la respuesta de la respiración de los microorganismos del suelo ante el contacto con las muestras de tejido vegetal senescentes provenientes de los maíces convencional e híbrido H-Ame15.
A partir del día 42, ocurrió la disminución de los niveles de CO2 en el grupo administrado con el maíz híbrido H-Ame15, cuyos valores igual que en el grupo expuesto a la glucosa, fueron inferiores en relación al grupo con el maíz convencional, conducta que se mantuvo hasta el día 63.
La utilización de la prueba de comparación permitió detectar diferencias entre los grupos evaluados, los días 5, 28 y 35 entre el control (glucosa) y con las muestras provenientes de ambos maíces, aquellos suplementados con los residuos vegetales tuvieron un efecto significativamente estimulante sobre los niveles de CO2 (p< 0,5). Desde el día 42 hasta el final del experimento las diferencias se localizaron entre el grupo en el que aconteció la exposición a los residuos de maíz convencional cuya respiración se incrementó con respecto al resto de los grupos (p< 0,5).
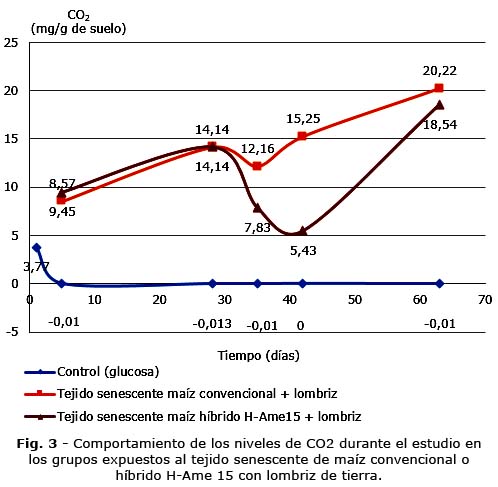
Los resultados de la cuantificación del CO2 en los grupos en los que además de suministrar el tejido vegetal se añadieron las lombrices de tierra se exponen en la figura 3.
Los resultados en el procesamiento estadístico mostraron diferencias significativas entre los conjuntos de datos entre el grupo control (glucosa) y los grupos expuestos a los restos de los maíces convencional e híbrido H-Ame15 (p< 0,5). La presencia de las lombrices estimuló la respiración de la comunidad microbiana (Fig. 2 y Fig. 3).
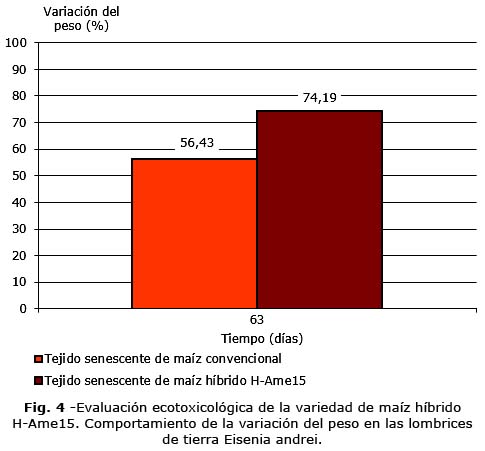
Las lombrices de tierra penetraron al sustrato en un tiempo inferior a una hora. No mostraron alteraciones anatómicas ni de su conducta. En la figura 4, se aprecia en ambos grupos un incremento del peso de los organismos al final del ensayo lo que se corresponde con la degradación de los residuos en estos grupos, el aumento fue mayor en el grupo expuesto al híbrido H-Ame15.
En los grupos en los que se incluyeron los restos de ambos maíces, se observó similitud en el patrón de descomposición con total desintegración el día 63, momento en que culminó el estudio.
DISCUSIÓN
El estado del suelo influye directamente en la producción agrícola, afecta la composición nutricional y los posibles residuos de químicos en los cultivos, lo cual puede tener consecuencias para la salud humana. Las modificaciones genéticas en el maíz transgénico, como el híbrido H-Ame15 buscan mejorar características agrícolas, pero pueden alterar el equilibrio ecológico, lo que a su vez puede impactar la calidad del cultivo y su inocuidad para el consumo humano.(15,16)
El efecto del maíz híbrido H-Ame15 en los ecosistemas del suelo es un indicador clave a estimar ya que refleja la estabilidad de los procesos ambientales, que de afectarse podría repercutir en la salud humana.(17,18,19)
Las funciones del suelo, como su calidad, dependen principalmente de factores bióticos, como la flora y la fauna, así como de su abundancia y composición. La fauna asociada abarca sus constituyentes y los procesos edáficos asociados, para mantener su aptitud. Esta consideración ambiental se centra entre otros, en los posibles efectos sobre su eficacia, los ciclos biogeoquímicos u otros procesos edáficos mediados por microbios cuando estos se ven afectados negativamente.(4) La mayoría de los estudios sugieren que las proteínas Bt de plantas transgénicas se descomponen rápidamente al penetrar a este compartimento y que no se bioacumulan.(20) No obstante, es necesario conocer cómo pueden incidir en la producción de CO2 en el suelo, que se genera mediante procesos biológicos, la descomposición de materia orgánica, la respiración de los organismos que lo habitan y los sistemas radiculares de las plantas.(21,22)
El pH del suelo es una propiedad química considerada como indicador de la disponibilidad de nutrientes al mostrar que tan ácida o alcalina resulta. Tal es su importancia que la mayoría de los microorganismos no sobreviven a pH inferiores a tres, ni superiores a 11.(23) En el caso de los grupos que contenían lombrices el pH inferior a 8,5 garantizó el bienestar de este invertebrado que se desarrolla de manera óptima con rangos de pH de 6,5 a 8,5.(24,25) El proceso de liberación de CO2, a través de la respiración edáfica, ocurre a 10 cm de profundidad en el suelo, donde se concentran los microorganismos involucrados en este proceso: Hongos y bacterias nitrificantes. Las variaciones por unidad de tiempo en la actividad respiratoria pueden ser interpretadas como reflejo de la actividad metabólica de los microorganismos que residen en determinado hábitat y permite conocer sobre la dinámica de su biota.(26,27)
Los microorganismos son capaces de degradar desde azúcares simples, proteínas, ácidos orgánicos hasta substratos muy complejos compuestos por hemicelulosas. Durante el proceso de degradación se conoce que en principio se desarrollan preferentemente grupos de bacterias no esporógenas, que utilizan los materiales orgánicos más asequibles, monosacáridos, disacáridos, aminoácidos y proteínas.(28)
En consonancia con los resultados, las poblaciones de bacterias heterotrópicas no mostraron afectación al ser expuestas a diferentes concentraciones de proteínas insecticidas provenientes de B. thuringiensis.(29)
De igual manera, varias investigaciones sobre el impacto de los cultivos Bt en los microorganismos del suelo no reportaron efectos significativos en experimentos de laboratorio, microcosmos y condiciones de campo.(30,31,32)
Asimismo, numerosos estudios publicados indican que la exposición a las proteínas Bt producida en plantas con protección incorporada no afecta negativamente a los microorganismos del suelo.(10,26) Además, las toxinas de Bacillus thuringiensis liberadas por los exudados radiculares y la biomasa del maíz Bt no tiene un efecto aparente sobre lombrices, nematodos, protozoos, bacterias ni hongos del suelo.(33,34)
Los resultados sugieren que el cultivo de maíz híbrido H-Ame15 disminuyó la actividad respiratoria de los microorganismos del suelo, lo que indica que podría alterar la dinámica de la comunidad microbiana del suelo, su funcionabilidad y diversidad.(35,36)
En el proceso de descomposición existen tres componentes importantes en la planta (celulosa, hemicelulosa y lignina), de ellos la lignina es el segundo en importancia es el compuesto aromático más abundante en la naturaleza y probablemente el más resistente a la degradación, consistente con sus funciones de brindar rigidez a las plantas vasculares y proteger los polisacáridos estructurales (celulosa y hemicelulosa) de las enzimas degradativas.(37) Algunos autores reportan una mayor concentración de lignina en cultivos Bt con respecto a los convencionales,(38) lo que puede explicar el comportamiento obtenido en el híbrido.
En estudios previos se analizó el impacto en los ecosistemas edáficos de las líneas de maíz con los eventos de transformación Bt 11 y Bt 176, que expresan el gen Cry1Ab de B. thuringiensis. Los residuos vegetales de plantas transgénicas, afectaron la respiración del suelo, las comunidades bacterianas y el establecimiento de micorrizas por endófitos autóctonos. Al igual que en la evaluación del efecto del maíz híbrido H-Ame15, se detectaron niveles de CO2, significativamente menores en las muestras de suelo que contenían residuos de maíz Bt.(39)
Estos resultados difieren con los obtenidos por otros autores, quienes demostraron que los híbridos modificados nunca fueron diferentes de sus correspondientes isolíneas en cuanto a la emisión de CO2.(40) Otras investigaciones concluyen que, si bien la presencia de tejido vegetal transgénico en el suelo puede producir un aumento y cambios transitorios en las poblaciones microbianas, sin atribuir efectos adversos a las proteínas Bt, así como la semejanza del tiempo de degradación de las proteínas insecticidas Cry y Vip3.(41)
En las variedades de maíz Bt Novelis y Valmont, que expresan la toxina Cry1Ab, los residuos de tallos y hojas tienen concentraciones respectivas de 3,9 y 0,8 ppm de peso seco. Los tratamientos con las variedades de maíz Bt mostraron un pico inicial significativo en la respiración de 0,7 mg de CO2/g de suelo. Al analizar la respiración de la comunidad microbiana del suelo luego del contacto con las proteínas Cry1Fa y Vip3A, a las respectivas concentraciones de 0,89 y 1,735 ppm de suelo, a través de los residuos del maíz híbrido H-Ame15, se obtuvieron niveles iniciales de CO2 de 7,61 mg/g de suelo.(42)
En los sistemas agrícolas, la biodiversidad del suelo es importante para sustentar los servicios ecosistémicos terrestres como la productividad, la mineralización de la materia orgánica, el reciclaje y disponibilidad de nutrientes, así como la formación de la estructura del suelo, entre otros. La biodiversidad del suelo depende de la macrofauna y la microfauna.(43,44)
Cada vez es mayor la evidencia del rol que desempeñan las lombrices de tierra y sus microbiomas intestinales en el mantenimiento del metabolismo de las lombrices, en la transformación de nutrientes, además de los compuestos exógenos en los suelos.(45,46,47) La presencia de las lombrices de tierra en el estudio de respiración de la comunidad microbiana del suelo proporciona una representación más holística de la dinámica del ecosistema ya que estas interactúan como puente entre diferentes niveles de fauna, lo que permite una comprensión más cercana a la realidad ecológica donde coexisten diversas formas de vida.(48,49)
Es necesario conocer si los cultivos Bt modifican los patrones de descomposición con respecto a los cultivos convencionales dado el impacto que puede tener en la disponibilidad de nutrientes y acumulación de biomasa vegetal en situaciones donde ocurra un enlentecimiento de la descomposición. Durante el proceso de descomposición de la hojarasca primero se libera la fracción lábil (azúcares y proteínas) y después la fracción recalcitrante, de más lenta descomposición, como las ligninas y los fenoles.(28)
Zhou X y otros(50) compararon los impactos de los restos del maíz Bt IE09S034 (Cry1Ie) con la línea isogénica Zong31, en la comunidad bacteriana y la actividad metabólica microbiana. Los resultados mostraron que los residuos de maíz Bt no tuvieron un impacto directo en la diversidad funcional microbiana y la actividad metabólica del suelo, a diferencia del entorno y el tiempo de descomposición.
En contraste a los hallazgos relacionados con la degradación del híbrido H-Ame15 y su variante convencional, Flores S y otros(15) demostraron que la degradación de las plantas Bt en el suelo fue más lenta que la de las plantas no Bt, resultados que lo relacionaron con el contenido de lignina que fue significativamente superior en el maíz Bt.(51)
Los riesgos ambientales que representan las toxinas Bt liberadas a través de los exudados radiculares de los cultivos transgénicos constituye una preocupación de la comunidad científica internacional.(52,53) Las plantas transgénicas contienen nuevos genes responsables de la expresión de cualidades que incrementan su capacidad de supervivencia y sus habilidades competitivas. Cada vez surgen más razones de índole ecológica que cuestionan los riesgos de estos cultivos el flujo de genes a organismos nativos, el desarrollo de resistencias en plagas diana, consecuencias directas e indirectas sobre organismos no diana y por lo tanto sobre el ecosistema.(54,55)
Los indicadores para la comprensión integral del impacto de los cultivos Bt en el suelo incluyen análisis de la diversidad, composición y función, dentro de esta última la respiración microbiana constituye un punto de partida útil.(56,57,58,59,60)
El maíz híbrido H-Ame15 no afectó los indicadores de la salud edáfica.
REFERENCIAS BIBLIOGRÁFICAS
1. Vidal Ledo MJ, Armenteros Vera I, Aparicio Suárez JL, Morales Suárez I, Portuondo Sao M. Una Salud [Internet]. Educación Médica Superior. 2021; 35(2):e2890. http://scielo.sld.cu/scielo.php?pid=S0864-21412021000200018&script=sci_arttext
2. Organisation for Economic Co-operation and Development, Safety Assessment of Transgenic Organisms in the Environment, Volume 10: OECD Consensus Document on Environmental Considerations for the Release of Transgenic Plants, Harmonisation of Regulatory Oversight in Biotechnology [Internet]. Paris: OECD; 2023. DOI: 10.1787/62ed0e04-en
3. Hilbeck A, Meyer H, Wynne B, Millstone E. GMO regulations and their interpretation: how EFSA’s guidance on risk assessments of GMOs is bound to fail [Internet]. Environ Sci Eur. 2020; 32:54. DOI: 10.1186/s12302-020-00325-6
4. Gallardo L. La biotecnología alimentaria: mitos, realidades y derecho. [Internet]. Corrientes: Editorial Moglia Ediciones. Corrientes; 2020. [acceso: 09/05/2023]. Disponible en:https://repositorio.unne.edu.ar/handle/123456789/29670
5. López Baroni M. Las tres Europas ante la encrucijada genómica [Internet]. Rev Bio Der. 2019. [acceso: 01/05/2023]; 47:77-92. Disponible en: http://www.mag.go.cr/rev_meso/v31n01_209.pdf
6. O´Farril LC. Transgénesis: una aproximación a sus riesgos y beneficios [Internet]. Acta méd centro. 2021 [acceso: 30/04/2023]; 15(1):[aprox. 13 pant.]. Disponible en: https://revactamedicacentro.sld.cu/index.php/amc/article/view/1239
7. Castro-Landin AL, Mayra Lisette ML, Luisa Anabel LA. El rol de los microorganismos en la fertilidad del suelo agrícola basado en una revisión de estudios recientes[Internet]. ISJ. 2023 [acceso: 18/05/2025]; 1(1): 26-37. Disponible en: https://innovasciencejournal.omeditorial.com/index.php/home/article/view/8
8. Téllez P. Obtención de cultivares transgénicos de maíz, resistentes a Spodoptera frugiperda y a glufosinato de amonio, alternativa para el alto rendimiento y conservación de insectos asociados. [Tesis Doctoral]. La Habana: Universidad Agraria; 2024. p. 204.
9. Centro Nacional de Toxicología. Instrucción al cliente maíz híbrido H-AME15. Serie de Instrucción al cliente: 07.La Habana: CENATOX; 2020.
10. Liu X, Liu S, Bai S, He K, Zhang Y, Dong H, et al. Toxicity of Cry- and Vip3Aa-Class Proteins and Their Interactions against Spodoptera frugiperda (Lepidoptera: Noctuidae) [Internet]. Toxins.2024; 16(4):193. DOI: 10.3390/toxins16040193
11. Thompson H, Elston Ch. What can laboratory studies tell us about potential effects of pesticides on non target arthropods populations and communities in the field? Integrated Environmental Assessment and Management [Internet]. 2024; 20(6): 2326–33. DOI: 10.1002/ieam.4987
12. FAO, ITPS, GSBI, CDB y CE. Estado del conocimiento sobre la biodiversidad del suelo - Situación, desafíos y potencialidades. Resumen para los formuladores de políticas [Internet]. Roma: FAO; 2021. DOI: 10.4060/cb1929es
13. Office of Chemical Safety and Pollution Prevention. OCSPP Ecological Effects Test Guidelines: Soil Microbial Community Toxicity Test [Internet]. OCSPP Guidelines 850.3200. United States: OCSPP; 2012. [acceso: 10/05/2012]. Disponible en: https://www.regulations.gov/document/EPA-HQ-OPPT-2009-0154-0020
14. IBM. SPSS. Versión 29 [software]. 2022. [acceso: 14/10/2023]. Disponible en: https://www.ibm.com/products/spss-statistics
15. Medrano Macías J, Morales Espinoza I, Benavides Mendoza A. Suelos y nutrición humana -énfasis en I, Se, Zn y Fe- [Internet]. Agraria. 2023; 20(3):17-2. Disponible en: https://www.revistaagraria.com/index.php/agraria/article/view/36
16. Liceaga Mendoza RI. Ruptura metabólica, (des)conocimiento ecológico y el cuerpo de los cuerpos: una aproximación a la continuidad entre la alimentación, salud y ambiente y su importancia política [Internet]. Argumentos. Revista de crítica social. 2023; 28: 417-447. Disponible en:https://dialnet.unirioja.es/servlet/articulo?codigo=9169846
17. Anand U, Vaishnav A, Sharma SK, Sahu J, Ahmad S, Sunita K, et al. Current advances and research prospects for agricultural and industrial uses of microbial strains available in world collections [Internet]. Sci Total Environ. 2022; 10(842): 1-22. DOI: 10.1016/j.scitotenv.2022.156641
18. Koller F, Schulz M, Juhas M, Bauer Panskus A. Then C. The need for assessment of risk sarising from interactions between NGT organisms from an EU perspective [Internet]. Environ Sci Eur. 2023; 35(27): 1-15. DOI:10.1186/s12302-023-00734-3
19. Caradus, JR. Intended and unintendedconsequences of geneticallymodifiedcrops – myth, fact and/ormanageableoutcomes? New Zealand Journal of Agricultural Research [Internet].2022; 66(6): 519–619. DOI: 10.1080/00288233.2022.2141273
20. Zurbrügg C, Hönemann L, Meissle M, Romeis J, Nentwig W. Decomposition dynamics and structural plant components of genetically modified Bt maize leaves do not differ from leaves of conventional hybrids.2010; 19(2):257-67. Disponible en: https://pubmed.ncbi.nlm.nih.gov/19609704/
21. Gomes DS, Barbosa AS, Santos TM, Santos SK, Silva JHCS, Aquino ÍS. Cinética de liberação de CO2, e decomposição da fitomassaem sistemas de uso e manejo do solo [Internet]. Research, Society and Development. 2021; 10(1): 1-14. DOI: 10.33448/rsd-v10i1.11413
22. Gomes DS, Santos SK, Silva JHCS, Medeiros T, Santos EDVS, Barbosa AS. CO2 flux e temperatura da superfície edáfica em áreas de caatinga [Internet]. Revista Brasileira de Geografia Física. 2021 [acceso: 04/11/2023]; 14(04):1898- 1908. Disponible en: https://periodicos.ufpe.br/revistas/index.php/rbgfe/article/view/248853
23. Valle MF, Castellari C, Andreoli Y, Diaz Delfino A, Picone L. Abundancia y distribución de la microbiota modificada por el uso del suelo en el sudeste bonaerense [Internet]. Agronomía & Ambiente. Revista de la Facultad de Agronomía (UBA). 2022 [acceso: 11/10/2024]; 42(1): 15-25. Disponible en: http://agronomiayambiente.agro.uba.ar/index.php/AyA/article/view/150/172
24. Machado Cuellar L, Rodríguez Suárez L, Murcia Torrejano V, Orduz Tovar SA, Ordoñez Espinosa CM, Suárez JC. Macrofauna del suelo y condiciones edafo climáticas en un gradiente altitudinal de zonas cafeteras, Huila, Colombia [Internet]. Rev. biol. Trop. 2021; 69(1): 102–12.DOI: 10.15517/rbt.v69i1.42955
25. Rincones PA, Zapata JE, Figueroa OA, Parra C. Evaluación de sustratos sobre los parámetros productivos de la lombriz roja californiana (Eisenia fetida) [Internet]. Información Tecnológica. 2023; 34(2):11-20. DOI: 10.4067/S0718-07642023000200011
26. Da Silva Gomes D, de Medeiros Santos T, da Silva Barbosa A, Kelly dos Santos S, de Sousa Melo T, de Souza Aquino. Effects of land-use changes on soil respiration [Internet]. Journal of Environmental Analysis and Progress.2022 [acceso: 05/03/2022]; 07(01): 9-15. Disponible en: https://www.researchgate.net/publication/357944605_Edaphic_breath_in_soil_use_and_management_systems
27. Mandal A, Sarkar B, Owens G, Thakur JK, Manna MC, Niazi NK, et al. Impact of genetically modified crops on rhizosphere microorganisms and processes: A review focusing on Btcotton [Internet]. Appl. Soil Ecol. 2020; 148:103492. DOI:10.1016/j.apsoil.2019.103492
28. Sánchez S, Crespo G, Hernández M, García Y. Factores bióticos y abióticos que influyen en la descomposición de la hojarasca en pastizales [Internet]. Pastos y Forrajes. 2008 [acceso: 13/09/2022]; 31(2):99-118. Disponible en: http://scielo.sld.cu/scielo.php?script=sci_arttext&pid=S0864-03942008000200001
29. Ferreira LHPL, Molina JC, Brasil C, Andrade G. Evaluation of Bacillus thuringiensis bioinsecticidal protein effects on soil microorganisms [Internet]. Plant and Soil. 2003; 256: 161–8. DOI: 10.1023/A:1026256700237
30. Oliveira AP, Pampulha ME, Bennett JP. A two-year field study with transgenic Bacillus thuringiensis maize: Effects on soil microorganisms [Internet]. Science of the Total Environment. 2008; 405:351-7. DOI: 10.1016/j.scitotenv.2008.05.046
31. Miethling-Graff R, Dockhorn S, Tebbe CC. Release of the recombinant Cry3Bb1 protein of Bt maize MON88017 into field soil and detection of effects on the diversity of rhizosphere bacteria [Internet]. Eur. J. Soil Biol. 2010 [acceso: 12/05/2025]; 46(1): 41–8. Disponible en: https://www.sciencedirect.com/science/article/abs/pii/S1164556309000995
32. Tan FX, Wang JW, Feng YJ, Chi GL, Kong HL, Qiu HF, et al. Bt corn plants and their straw have no apparent impact on soil microbial communities [Internet]. Plant Soil. 2010; 329: 349–64. DOI:10.1007/s11104-009-0163-2
33. Sanvido O, Romeis J, Bigler F. Ecological impacts of genetically modified crops: ten years of field research and commercial cultivation [Internet]. Advances in Biochemical Engineering and Biotechnology. 2007;107:235 –78. DOI: 10.1007/10_2007_048
34. Saxena D, Stotzky G. Bacillus thuringiensis (Bt) toxin released from root exudates and biomass of Btcorn has no apparent effect on earthworms, nematodes, protozoa, bacteria, and fungi in soil [Internet]. Soil Biology and Biochemistry.2001; 33(9): 1225–30. DOI: 10.1016/S0038-0717(01)00027-X
35. Chassain J, VieubléGonod L, Chenu C, Joimel S. Role of different izeclasses of organisms in croppedsoils: What do litter bag experiments tellus? A meta-analysis [Internet]. Soil Biology and Biochemistry. 2021; 162. DOI: 10.1016/j.soilbio.2021.108394
36. Castellanos Gonzalez L, Capacho Mogollón AE, Castellanos Hernández L. Variaciones de la microfauna del suelo con la implantación de 18 modelos agroecológicos en 6 municipios de Norte de Santander, Colombia [Internet]. Inge Cuc. 2020 [acceso: 10/05/2025]; 17(1): 81-95. Disponible en: https://revistascientificas.cuc.edu.co/ingecuc/article/view/3078
37. Sanclemente Reyes ÓE, Ararat Orozco MC, Mejía Ballesteros JE, Ríos LÁ, Montoya Rendón JC, Arango Arias ÁM, Arteaga Aguilera LF, et al. Relaciones agroambientales en sistemas productivos rurales [Internet]. 1 ed. Bogotá: Sello Editorial UNAD; 2021. DOI: 10.22490/9789586518048
38. Gómez Lende S. Cultivos transgénicos y acumulación por desposesión en Argentina (1991-2021): Categorías de análisis y ejemplos empíricos [Internet]. Clivajes. 2024 [acceso: 17/02/2025]; 17:102-33. Disponible en: https://clivajes.uv.mx/index.php/Clivajes/article/view/2793/4542
39. Castaldini M, Turrini A, Sbrana C, Benedetti A, Marchionni M, Mocali S, et al. Impact of Bt Corn on Rhizospheric and Soil Eubacterial Communities and on Beneficial Mycorrhizal Symbiosis in Experimental Microcosms [Internet]. Appl. Environ. Microbiol. 2005; 71(11): 6719–29. DOI: 10.1128/AEM.71.11.6719-6729.2005
40. Zurbrügg C, Hönemann L, Meissle M, Romeis J, Nentwig W. Decomposition dynamics and structural plant components of genetically modified Bt maize leaves do not differ from leaves of conventional hybrids [Internet]. Transgenic Res. 2010; 19: 257–67. DOI:10.1007/s11248-009-9304-x
41. Ombudsman, Biopesticides and Pollution Prevention Division (7511P). Bacillus thuringiensis Vip3Aa20 Insecticidal Protein and the Genetic Material Necessary for Its Production (via Elements of VectorpNOV1300) in Event MIR162 Maize (OECD) Unique Identifier: SYN-IR162-4) (006599) Fact Sheet [Internet].Washington DC: Office of Pesticide Programs Environmental Protection Agency; 2008.[acceso: 17/02/2025]. Disponible en: https://www3.epa.gov/pesticides/chem_search/reg_actions/registration/fs_PC-006599_26-Dec-08.pdf
42. Raubuch M, Roose K, Warnstorff K, Wichern F, Joergens en R. Respiration pattern and microbial use of field-grown transgenic Bt-maize residues [Internet]. Soil Biology and Biochemistry. 2007; 39(9):2380-9. DOI:10.1016/j.soilbio.2007.04.012
43. Chen XD, Dunfeld KE, Fraser TD, Wakelin SA, Richardson AE, Condron LM. Soil biodiversity and biogeo chemical function in managed ecosystems [Internet]. Soil Research 2020; 58: 1–20. DOI: 10.1071/SR19067
44. Insfrán Ortiz A, Rey Benayas JM, Cayuela Delgado L. Agroforestry improves soil fauna abundance and composition in the Atlantic Forest of Paraguay[Internet]. Agroforest Syst. 2023; 97:1447–63. DOI: 10.1007/s10457-023-00869-5
45. Liu P, Yang Y, Li M. Responses of soil and earthworm gut bacterial communities to heavy metal contamination [Internet]. Environmental Pollution. 2020; 265 (Part B). DOI:10.1016/j.envpol.2020.114921
46. Sun M, Chao H, Zheng X, DengSh, Ye M, Hu F. Ecological role of earthworm intestinal bacteria in terrestrial environments: A review [Internet]. Science of The Total Environment. 2020: 740. DOI: 10.1016/j.scitotenv.2020.140008
47. Ortíz-Gamino D, Gregorio J, Barois I. ¿Las lombrices de tierra tienen potencial biotecnológico? [Internet]. Elementos 123. 2021 [acceso: 24/01/2025]; 71-7. Disponible en: https://elementos.buap.mx/directus/storage/uploads/00000006199.pdf
48. Ahmad A, Aslam Z, Bellitürk K, Iqbal N, Idrees M, Nawaz M, et al.Earthworms and vermicomposting: A review on thestory of blackgold [Internet]. Journal of InnovativeSciences 2021; 7(1):167-73. DOI: 10.17582/journal.jis/2021/7.1.167.173
49. Wang L, Fan Y, Zou L, Ge L, Jiang W, Chao S, et al. Bt toxins alter bacterial communities and their potential functions in earthworm intestines [Internet]. Environmental Pollution. 2025 [acceso: 18/05/2025]; 367: 125591. Disponible en: https://www.sciencedirect.com/science/article/abs/pii/S026974912402308X?via%3Dihub
50. Zhou X, Liang J, Luan Y, Song X, Zhang Zh. Characterisation of Bt maize IE09S034 in decomposition and response of soil bacterial communities [Internet]. Plant, Soil and Environment. 2021; 67(5): 286-98. DOI: 10.17221/629/2020-PSE
51. Flores S, Saxena D, Stotzky G. Transgenic Bt plants decompose less in soil than non-Bt plants [Internet]. Soil BiolBiochem.2005; 37(6): 1073–82. DOI: 10.1016/j.soilbio.2004.11.006
52. Oliveira-Filho EC, Grisolia CK. The Ecotoxicology of Microbial Insecticides and Their Toxins in Genetically Modified Crops: An Overview [Internet]. Int J Environ Res PublicHealth. 2022; 19(24): 1-14. DOI: 10.3390/ijerph192416495
53. Karim AA, Idris AB, Yilmaz S. Bacillus thuringiensis pesticidal toxins: A global analysis based on a scientometric study (1980-2021) [Internet]. Heliyon. 2023 [acceso: 04/07/2024]; 9(8): e18739. Disponible en:https://www.cell.com/action/showPdf?pii=S2405-8440%2823%2905938-8
54. Dunfield KE, Germida JJ. Impact of genetically modified crops on soil- and plant-associated microbial communities [Internet]. J Environ Qual. 2004; 33(3): 806-15. DOI: 10.2134/jeq2004.0806
55. Messéan A, Álvarez F, Devos Y, Camargo AM. Assessment of the 2021 post-market environmental monitoring report on the cultivation of genetically modified maize MON 810 in the EU [Internet]. EFSA J. 2023 [acceso: 10/05/2025]; 21(12):e8411. Disponible en: https://pmc.ncbi.nlm.nih.gov/articles/PMC10699111/
56. Tikhonova E, Shestibratov K. Assessing Impacts of Transgenic Plants on Soil Using Functional Indicators: Twenty Years of Research and Perspectives [Internet]. Plants. 2022; 11(18): 2439. DOI: 10.3390/plants11182439
57. Kochiieru M, Veršulienė A, Feiza V, Feizienė D. Trend for Soil CO2 Efflux in Grassland and Forest Land in Relation with Meteorological Conditions and Root Parameters [Internet]. Sustainability. 2023; 15(9): 7193. DOI: 10.3390/su15097193
58. Barrera León J, Barrezueta Unda S, García Bastidas RM. Evaluation of Soil Quality Indices of Different Crops Under Different Topographic Conditions [Internet]. Revista Metropolitana de Ciencias Aplicadas. 2020 [acceso: 18/05/2025]; 3(1): 182-90. Disponible en:https://www.researchgate.net/publication/340789690_EVALUATION_OF_SOIL_QUALITY_INDICES_OF_DIFFERENT_CROPS_UNDER_DIFFERENT_TOPOGRAPHIC_CONDITIONS
59. Morales-Rojas E, Chávez-Quintana S, Hurtado-Burga R, Milla-Pino M, Sanchez-Santillán TE. Martos Collazos-Silva. Macrofauna edáfica asociada al cultivo de maíz (Zea maíz) [Internet]. J. Selva Andina Biosph. 2021; 9(1): 15–24. DOI: 10.36610/j.jsab.2021.090100015
60. Vázquez C, de Goede RGM, Rutgers M, de KoeijermT J, Creamer RE. Assessing multifunctionality of agricultural soils: Reducing the biodiversity trade-off [Internet]. European Journal of Soil Science. 2020; 72(4):1624–39. DOI: 10.1111/ejss.13019
Conflictos de interés
Los autores no declaran conflictos de interés.
Financiamiento
El estudio lo financió el proyecto del CIGB para el desarrollo del maíz híbrido H-Ame15.
Contribuciones de los autores
Conceptualización: Odette Beiro Castro, Yordanka Dominguez Linares, Ana Cristina Noa Rodríguez.
Curación de datos: Licette Leon Barreras.
Análisis formal: Mario Pablo Estrada García.
Investigación: Odette Beiro Castro, Yordanka Dominguez Linares, Ana Cristina Noa Rodríguez.
Metodología: Odette Beiro Castro, Yordanka Dominguez Linares, Ana Cristina Noa Rodríguez.
Supervisión: Mario Pablo Estrada García.
Redacción - borrador original: Odette Beiro Castro, Yordanka Dominguez Linares, Aymee Ferrer Colás.
Redacción - revisión y edición: Odette Beiro Castro, Yordanka Dominguez Linares, Aymee Ferrer Colás, Pilar Telléz-Rodríguez.
Declaración de disponibilidad de datos
Todos los datos se recogen en el artículo.